
Note: A bit of a disclaimer is necessary. This will probably
be one of my most indulgent post series, as this is probably my most favorite
topic in paleontology. Walruses are a totally weird and fascinating group to
study, and I hope some of my enthusiasm for these fantastic blubbery beasts
shines through. For later entries in this series, see:
Walrus Evolution, part 2
Walrus Evolution, part 3
Walrus Evolution, part 4
Walrus Evolution, part 5
Introduction
The walrus (Odobenus rosmarus) is one of the most peculiar and charismatic of all modern mammals, and easily the most distinctive of all pinnipeds. The modern walrus is characterized by its great bulk and most obviously by its large tusks; currently the walrus is restricted to the Arctic region, inhabiting the coasts of Greenland, Baffin Island, and Northern Quebec in the Northwestern Atlantic region, Svalbard, Novaya Zemlya, the Barents Sea, and the Laptev Sea in the Arctic region, and the Chukchi Sea and Bering Sea around Alaska and the Anadyr Peninsula (Russia).
The modern walrus feeds predominantly on mollusks, and uniquely feeds by placing the shell inside its lips and sucking the muscle tissue directly out of the poor little clam: the shell never enters the mouth, and the teeth are not used at all during feeding. Instead, the highly vaulted palate of the walrus permits the muscular tongue to act like a powerful piston as it is retracted. It also allows the walrus to jet out water onto the seafloor to uncover individual mollusks during foraging. The walrus is otherwise unique amongst pinnipeds by routinely mating in the water; unlike many other pinnipeds, the walrus also is flexible when it comes to where it hauls out – it can haul out on icepack or rocky beaches. Many other pinnipeds only haul out on a certain substrate, whether its sand (e.g. NZ sea lion, elephant seals), rocky shore (NZ fur seal), or ice (most arctic and Antarctic true seals). Most famous of all, however, are the spectacular tusks of walruses – greatly enlarged upper canines that are so long they protrude eternally from the animal’s mouth. What did tusks evolve for? A few studies have advanced various hypotheses, and the evolution of tusks will be considered in a later post in this series.
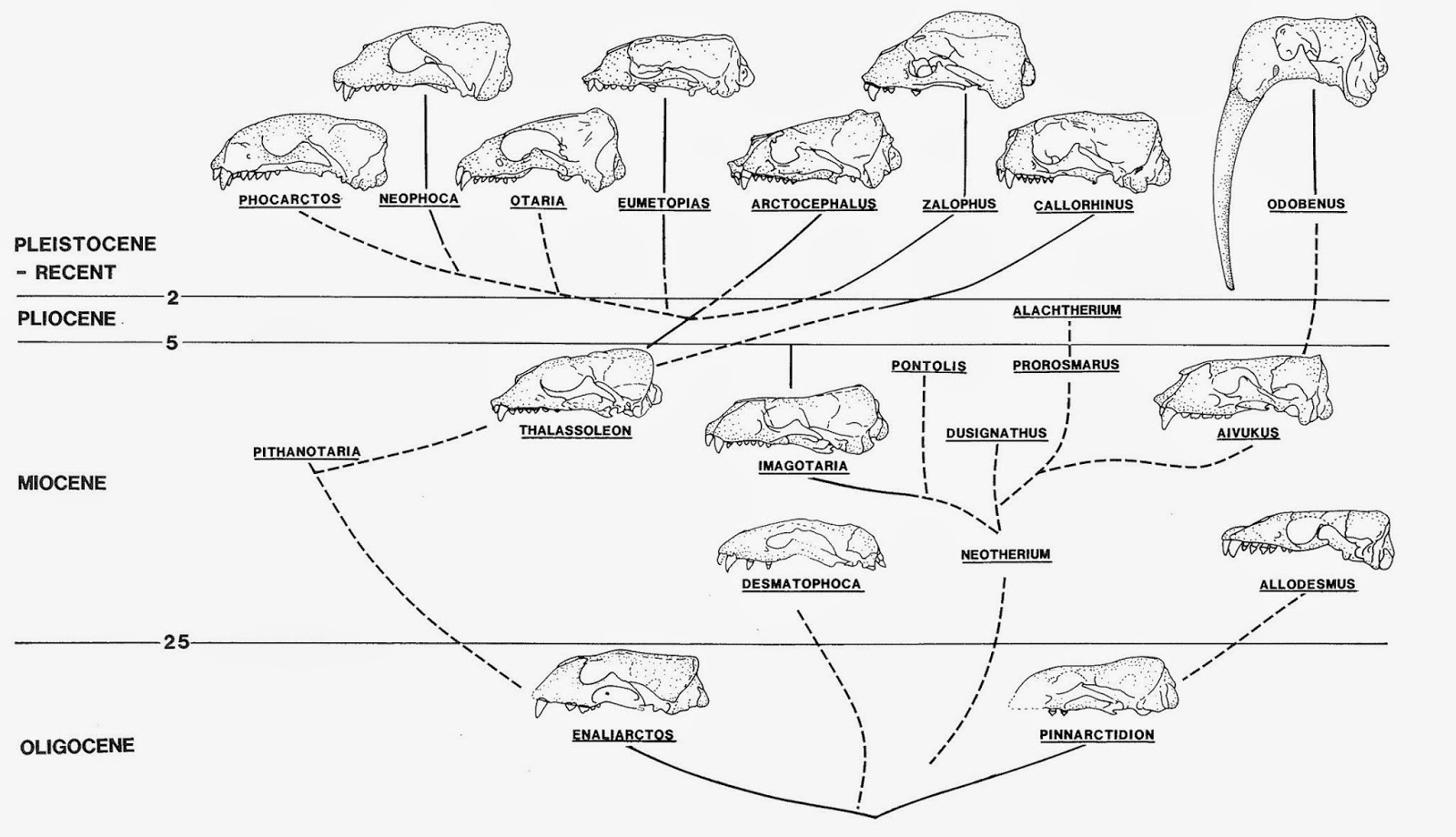
A somewhat outdated hypothesis of walrus relationships (from Barnes et al., 1985) - however, this figure is still informative, and is one of the only cladograms labeled with anatomical illustrations. I'm currently working on enough skull illustrations to make a modern version with an updated phylogeny. Notably absent from this phenogram are true seals, which were left out in the spirit of pinniped diphyly.

The most up-to-date hypothesis of walrus phylogeny, showing many of the interrelationships amongst the Odobenidae. For the purpose of this blog post, note the basal position of Prototaria, Proneotherium, Neotherium, and Kamtschatarctos. From Boessenecker and Churchill (2013).
Lastly, the phylogenetic relationships of walruses have proved contentious. Although the relationships within the family Odobenidae are fairly well resolved (compare the cladistic results of Deméré, 1994, Deméré and Berta, 2001, Kohno, 2006, and Boessenecker and Churchill, 2013), the relationships of the family to other pinniped clades has been a major point of debate, if not a source of ideological schism within pinniped paleontology (an admittedly small subdiscipline). Traditionally, walruses have been placed into a clade called Otarioidea including all pinniped groups except for the Phocidae; Otarioidea (or a much more taxonomically inclusive Otariidae of some authors, e.g. Mitchell and Barnes) would include the Otariidae, Odobenidae, Desmatophocidae, and the “enaliarctines” (early primitive pinnipeds). This traditional view has generally been based on non-cladistic interpretations of morphological evidence. Viewpoints typically endorsing monophyly of Otarioidea also consider the pinnipeds to be diphyletic, with phocids evolving from an as-yet unidentified mustelid ancestor and otarioids evolving from a bear-like ancestor. A more recent interpretation based on robust cladistic work (Berta and Wyss, 1994) endorses a phylogeny of pinnipeds that is fundamentally different in three ways: 1) pinnipeds are indeed monophyletic, 2) extant phocid seals and extinct desmatophocids (e.g. Allodesmus) form a clade, the Phocoidea, and 3) the walruses (Odobenidae) are sister to the Phocoidea, forming the clade Phocomorpha – including walruses, true seals, and desmatophocids. The Phocomorpha hypothesis is superior to the Otarioidea hypothesis by virtue of actually having cladistic evidence that supports it. Further complicating these matters is the fact that modern molecular studies – virtually all of them – unilaterally demonstrate that pinnipeds are a monophyletic group, typically with 100% branch support in every study. However, virtually all of these molecular studies also indicate – again with robust support – that otariids and odobenids form a monophyletic group to the exclusion of phocids. Ironically, for once, the molecular work doesn’t really go at odds with the morphological evidence so much as it actually provides sort of a satisfying compromise between two extremes. Regardless, the molecular hypotheses for relationships within pinnipeds are still lacking support from morphological evidence. More on this will be discussed later.
The rest of this post will focus on sea lion-like pinnipeds now known to be walruses, but they lack tusks and were small in stature and did not live on ice; how do we know they're walruses? A few unique features unite all fossil and modern walruses: an antorbital process of the rostrum composed of both frontal and maxilla, a thickened "pterygoid strut" at the posterior end of the palate, an internal ridge called the tentorium overlapping the inner ear bone, lingual cusps on the upper anterior premolar teeth, and a triple rooted upper molar (a reversal to the primitive condition in terrestrial arctoids, whereas in enaliarctines the tooth is double rooted; both dental features are lost in later odobenids). A few postcranial features (mostly humerus, radius, the first metacarpal, and a wrist bone called the scapholunar), also unite all walruses (some early walruses are not yet known from well preserved postcranial bones).

One of the earliest discovered fossils of a stem walrus: the syntype astragalus of Neotherium mirum, described by Kellogg (1931) from the middle Miocene Sharktooth Hill Bonebed.
The earliest walruses
The earliest known walruses evaded identification for nearly forty years. Part of the problem was that prior to the 1970’s, nobody knew what a primitive pinniped looked like, and the scarcity of early molecular work meant that nobody knew where pinnipeds fit within Carnivora, further meaning that nobody knew what a primitive pinniped should look like. Although now known to be an early walrus, the pinniped Neotherium mirum was named in 1931 by Remington Kellogg from the Sharktooth Hill Bonebed based on a paltry handful of non-associated ankle bones. Kellogg tentatively considered it to represent an otariid pinniped (and judging from the size of the postcranial bones accurately estimated its size to be somewhat smaller than an extant adult female California sea lion), but even later workers (e.g. Mitchell, 1961) considered the identity of Neotherium to be tenuously known at best. In 1968, Ed Mitchell described the new genus Imagotaria from the late Miocene Monterey/Sisquoc Formation of Santa Barbara County, which at the time he considered to be an ancestral sea lion closely related to the gigantic pinniped Pontolis magnus. Pontolis had been discovered at about the turn of the century in the Empire Formation near Coos Bay, Oregon, and until the 1990’s was only known from a gigantic incomplete braincase.
In 1973, Ed Mitchell and Richard Tedford described the early pinniped Enaliarctos mealsi from the Jewett Sand at Pyramid Hill, a prolific locality near Bakersfield, California (not too far away from the more famous Sharktooth Hill) which has yielded a lot of important marine vertebrate fossils from about the Oligo-Miocene boundary. Enaliarctos mealsi effectively resembles a fur seal with the dentition and other skull features of a terrestrial carnivore. While clearly not a walrus, at least by this point the primitive condition of the pinniped skull was known and could be compared with later fossils.
In 1977, a monograph was published by Charles Repenning and Richard Tedford titled “Otarioid Seals of the Neogene”, one of the most important contributions – if not the most important contribution - in the history of fossil pinniped studies. I affectionately call their paper the “pinniped bible”. In their paper, they described a new fossil walrus that will be discussed in more detail later, Aivukus cedrosensis – and described more skulls, mandibles, and postcranial bones of Imagotaria. These new discoveries indicated that in the past, walruses did not always have tusks and had a more generalized sea lion-like skull and dentition, probably signifying a more generalized ecology. Imagotaria and company will be discussed in more detail in the next post.

The indifferently preserved holotype skull and (somewhat better) mandible of Kamtschatarctos sinelnikovae, from Dubrovo (1981).
The first early walruses to have been described from skulls were not from North America, but rather from the western North Pacific – Kamtschatarctos sinelnikovae, from the middle Miocene of Kamtchatka (Dubrovo, 1981) in eastern Russia (then, the Soviet Union), and Prototaria primigena from the early middle Miocene of Japan (Takeyama and Ozawa, 1984). Neither of these were immediately recognized as walruses, but Barnes (1989) identified that both shared features with Imagotaria and (at the time) undescribed cranial material of Neotherium. Initially, Dubrovo (1981) named a new subfamily, the Kamtschatarctinae, to contain Kamtschatarctos (only) within the family “Enaliarctidae”. The Russian species is known only from a single squashed and incomplete skull with a rather delicate mandible. The most immediate alarm bell about this skull is its size – it’s quite a bit larger than all known enaliarctine material; so far as we know, all described enaliarctines were approximately the size of a harbor seal or so. Dubrovo (1981) drew attention to the root condition, although Barnes (1989) correctly indicated that the rooting was a primitive condition, and that the mandible (and root condition of the dentition) is quite similar to Imagotaria downsi in some regards. Despite being much more similar to Enaliarctos in overall skull morphology than Kamtschatarctos, Takeyama and Ozawa (1984) considered the much better preserved skull of Prototaria to be an early otariid (rather than an enaliarctine) morphologically intermediate between Enaliarctos and the fossil fur seal Thalassoleon. However, Barnes (1989) identified a number of primitive features it shares with undescribed cranial material of Neotherium, and even went so far as to reclassify it as Neotherium primigena.

The holotype skull and partial skeleton of Prototaria primigena, described by Takeyama and Ozawa (1984) from the early middle Miocene of Japan. Unfortunately, only the skull has been described and it is unclear if and when the postcrania will be described.
In 1994, Naoki Kohno described a second species of Prototaria from Japan, and named it Prototaria planicephala, alluding to its relatively flat skull. Both species of Prototaria are relatively tiny – about the size of a modern harbor seal – with relatively short snouts (rostra) and a generalized dentition. Both species are known from the early Middle Miocene of Japan. Kohno (1994) indicated that new skull material of Neotherium demonstrated that Prototaria was in fact a distinct, separate genus, primarily by the shorter rostrum, a more massively constructed zygomatic arch, and a few less interesting differences. Prototaria exhibits at least one feature (synapomorphy) uniting all walruses – a thickened pterygoid strut in the basicranium, a bony strut laterally adjacent to the internal choanae and posterior to the palate; this feature is present in most archaic walruses, differentiating them from all other pinnipeds – and is secondarily lost in later walruses. Other than this single important feature, Prototaria is nearly identical to enaliarctines like Enaliarctos and Pteronarctos, even primitively possessing small supraorbital processes (primitive for carnivorans, reduced in most odobenids, desmatophocids, and phocid seals).

Illustration of the holotype skull of Prototaria planicephala from the early middle Miocene of Japan, from Kohno (1994).
The next significant advance was the publication of skull material of Neotherium mirum and the naming of Proneotherium repenningi, from the middle Miocene Sharktooth Hill Bonebed and the late early-early Middle Miocene Astoria Formation of Oregon (respectively). Proneotherium is known by numerous skulls and a partial postcranial skeleton, and at least one mandible (Kohno et al., 1995; Deméré and Berta, 2001; Boessenecker and Churchill, 2013). Proneotherium has a more or less enaliarctine-like skull and postcranial skeleton, but already has at least one postcranial synapomorphy – a medial process on the calcaneum (the heel bone). Proneotherium also has an expanded pterygoid strut like Prototaria, and has a posterolingual shelf on some of its cheek teeth – a feature not preserved in Prototaria. A feature Morgan Churchill and I found in a referred mandible of Proneotherium which we figured in our paper on Pelagiarctos – is the position of the deepest part of the tooth-bearing part of the mandible (the “horizontal ramus”). In Enaliarctos and Pteronarctos, the deepest part is positioned at the lower molar, which is still shaped as a large carnassial shearing tooth; this is the primitive condition for carnivores. However, in Proneotherium, all later walruses, and modern sea lions and fur seals, the deepest part of the mandible is positioned anteriorly at the canines, giving the impression of a “deep” or “thick” chin. This condition is generally lacking in most modern and fossil true seals and the extinct pinniped Allodesmus, but is present in the other desmatophocid, Desmatophoca. Kohno et al. (1995) described the holotype skull and also designated a paratype skull; Deméré and Berta (2001) referred several additional skulls, including one associated with a partial postcranial skeleton. As an aside, Larry Barnes told me that Proneotherium was a pet name (since it was smaller, older, and thought to be ancestral to Neotherium) for this material used by the late Charles Repenning during the 1980’s; Barnes (in Kohno et al., 1995) decided to use the name and named the species after Repenning (a fitting honor for Rep). I never met Repenning unfortunately, as he was murdered in his home in 2005, the year I started marine mammal paleontology research.

Well-preserved skulls of Proneotherium repenningi from the early middle Miocene Astoria Formation of Oregon described by Demere and Berta (2001).
The last of the early small-bodied odobenids, Neotherium, is known widely from the Sharktooth Hill Bonebed. It was first described by Kellogg (1931) based upon a handful of non-associated cranial elements. Subsequent workers referred various bits including a humerus (Mitchell, 1961), radius (Repenning and Tedford, 1977), a mandible (Barnes, 1988) and finally a complete skull (Kohno et al., 1995). Repenning and Tedford (1977) noted that material of Neotherium effectively looked like miniature bones of Imagotaria; because only one small bodied walrus is known from the Sharktooth Hill Bonebed, isolated bones matching the size are referable to the species if they look similar to Imagotaria. On the other hand, Imagotaria is known from numerous specimens with associated craniodental and postcranial elements, whereas all material of Neotherium from the bonebed is isolated and not associated. Neotherium is notably distinct from Proneotherium, Prototaria, and enaliarctines in its complete lack of supraorbital processes – the first walrus to have lost these bumps above the orbits.

Comparison of the skulls of Proneotherium repenningi (left), Neotherium mirum (middle), and Imagotaria downsi (right).
In summary – during the early and middle Miocene, we have the earliest walruses evolve from enaliarctines. These early diverging walruses were small, probably about the same size as a harbor seal – or a small sea lion in the case of Neotherium. The dentition of these walruses was barely different from enaliarctines, although they lacked the dedicated carnassial shearing teeth of enaliarctines. Mitchell and Tedford (1973) suggested that Enaliarctos may have needed to return to land in order to chew, given that it was still using a primitive shearing dentition (some in-preparation work by Morgan Churchill will be weighing in on this in more detail). If this assertion is true, then the earliest walruses were certainly already pierce or raptorial feeders – biting prey items and swallowing them whole. Importantly, the palates of these early walruses were relatively flat – indicating that suction of prey into the mouth was not yet an important aspect of their feeding ecology (similar to most true seals and otariids).

A well-preserved skull of Neotherium mirum from the middle Miocene Sharktooth Hill Bonebed described by Kohno et al. (1995).
One curious trend seen in early walruses is the process of “molarization”. In these early walruses, the anterior premolars begin to look more and more like the molars, developing a thickened lingual cingulum (lingual = tongue side of the tooth, and cingulum is a ridge of enamel along the base of the crown) and even developing a thickened posterior root (in enaliarctines, anterior premolars have two skinny, cylindrical roots). Usually a transversely thickened root suggests that two roots have been fused into one; some of the upper teeth (notably the upper molar and upper fourth premolar in caniform carnivores) are primitively triple-rooted, with one anterior root and two posterior roots; one way to fuse up these roots during the process of tooth simplification is to fuse the two posterior roots together. This leads to a tooth that is double-rooted, but has a small cylindrical anterior root and a much wider posterior root; this condition is then typically reduced further into two cylindrical roots (as in most modern true seals), and eventually into a single fused root (as in the modern walrus and extant fur seals and sea lions). Tooth rooting can be a good “barometer” of how primitive or derived a particular fossil pinniped is, as the dentition has been simplified in all groups from the ancestral enaliarctine condition – and single-rooted teeth have evolved no fewer than six or seven times amongst pinnipeds (Boessenecker, 2011). Early walruses, on the other hand, seemed to be doing the opposite: the anterior premolars were being increased in size and complexity, adding a thickened lingual cingulum and even developing small cusps on it. It’s not really clear why this happened, or what the advantage was – the general rule of dental evolution in pinnipeds (and marine mammals in general) is tooth simplification. Only a few groups of marine mammals ever added any cusps to their teeth: basilosaurids (and some toothed mysticetes) added a bunch of cusps to their cheek teeth, as did leopard and crabeater seals. Other marine mammals such as Odobenocetops, narwhals (Monodon), and later walruses increased the size of some teeth and developed tusks, but I imagine that’s something much more easily governed by rampant sexual selection (again, more on that in a later post).
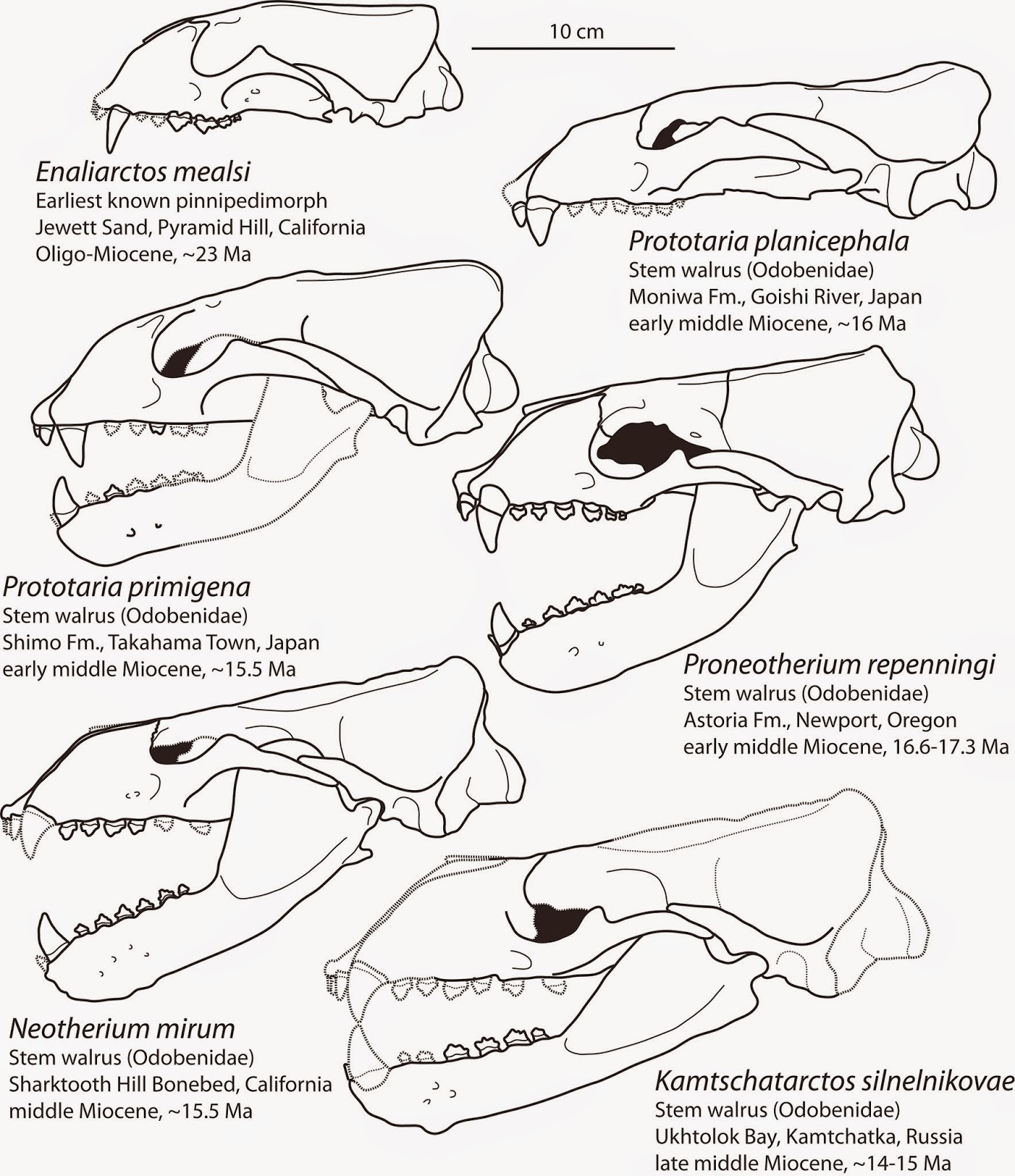
Line drawings of skulls in lateral view, including Enaliarctos mealsi (upper left) and Prototaria, Proneotherium, Neotherium, and Kamtschatarctos.
Postcranial bones have only really been published for Proneotherium (Deméré and Berta, 2001), although a handful of comparable elements have been published for Neotherium; they do not strongly differ, and functional statements/interpretations based on Proneotherium are likely to apply equally to Neotherium. The preserved hindlimb of Proneotherium shows it had various hallmark features of modern pinnipeds (Deméré and Berta, 2001), including a short femur and long tibia, long and robust lateral and medial digits (digits I and V) of the hindflipper with shorter intermediate digits (e.g. digits II, III, and IV). These two osteological features indicate that Proneotherium – and likely most other early walruses – already had a permanently flexed knee and also fan-shaped symmetrical hindflippers. However, Proneotherium primitively retains a pit on the head of the femur for the teres ligament (a ligament that keeps the head of the femur in the hip socket, missing in modern pinnipeds – both the ligament and the pit), and also primitively retains a lesser trochanter (a bump below the head of the femur, missing in many modern pinnipeds). Lastly, the ankle of Proneotherium is rigidly constructed indicating a substantial amount of terrestrial locomotion, as in modern otariids and terrestrial carnivores; the ankle of phocid seals is constructed as a loose ball-and-socket joint, and odobenine walruses have an intermediate but nonetheless loose ankle joint. This rigid ankle would have constrained movement of the hindflipper into flexion and extension and reducing rotation of the ankle – a retained adaptation for terrestrial walking.
This summary has highlighted a primitive pinniped that doesn’t really recall a mental image that looks anything like a walrus. Why is that? Pinnipeds began to diversify shortly after their invasion of the marine realm, and walruses are the earliest modern family of pinniped for which we can assign any fossils (specifically, Prototaria from Japan). The oldest known true seals are a bit younger (~15 Ma, Leptophoca from the Calvert Formation of Maryland; although the fragmentary Afrophoca is somewhat older, ~19 Ma), and the oldest otariids are a lot younger (~8-10 Ma, Pithanotaria from the Monterey Formation and Santa Margarita Sandstone of California). Although being relatively similar to primitive enaliarctines in most respects, these early walruses have just enough skull features to pull them closer to the modern walrus in cladistic analyses. Sometimes these early diverging walruses have been assigned to the subfamily “Imagotariinae” – but the “Imagotariinae” is not monophyletic (because it includes only primitive species and none of the later odobenids). I’ve only outlined the small-bodied early “imagotariines” here; the next post will cover the larger imagotariines like Imagotaria, Pelagiarctos, and Pseudotaria. At least three more posts will follow after that: a post on the bizarre dusignathine walruses, odobenine walruses and their evolutionary biogeography, and another post on the evolution of tusks.
References
Barnes, L.G. 1988. A new fossil pinniped (Mammalia: Otariidae) from the middle Miocene Sharktooth Hill Bonebed, California. Contributions in Science 396:1-11
Barnes LG, Domning DP, Ray CE. 1985. Status of studies on fossil marine mammals. Marine Mammal Science 1:15-53.
Boessenecker, R.W. 2011. New records of the fur seal Callorhinus (Carnivora: Otariidae) from the Plio-Pleistocene Rio Dell Formation of Northern California and comments on otariid dental evolution. Journal of Vertebrate Paleontology 31(2):454-467.
Boessenecker, R.W., and M. Churchill. 2013. A Reevaluation of the Morphology, Paleoecology, and Phylogenetic Relationships of the Enigmatic Walrus Pelagiarctos. PLoS One 8(1):e5411
Deméré. 1994. The Family Odobenidae: A phylogenetic analysis of fossil and living taxa. Proceedings of the San Diego Society of Natural History 29:99-123
Deméré, T.A., and A. Berta. 2001. A re-evaluation of Proneotherium repenningi from the middle Miocene Astoria Formation of Oregon and its position as a basal odobenid (Pinnipedia: Mammalia). Journal of Vertebrate Paleontology 21(2): 279-310.
Dubrovo, I.A. 1981. A new subfamily of fossil seals (Pinnipedia, Kamtschatarctinae subfam. nov.). Doklady Earth Science Sections 256:202-206
Kellogg, R. 1931. Pelagic mammals of the Temblor Formation of the Kern River region, California. Proceedings of the California Academy of Science 19(12):217-397.
Kohno, N. 1994. A new miocene pinniped in the genus Prototaria (Carnivora: Odobenidae) from the Moniwa Formation, Miyagi, Japan. Journal of Vertebrate Paleontology 14(3):414-426
Kohno, N. 2006. A new Miocene odobenid (Mammalia: Carnivora) from Hokkaido, Japan, and its implications for odobenid phylogeny. Journal of Vertebrate Paleontology 26(2):411-421
Kohno, N., L. G. Barnes, and K. Hirota. 1995. Miocene fossil pinnipeds of the genera Prototaria and Neotherium (Carnivora; Otariidae; Imagotariinae) in the North Pacific Ocean: Evolution, relationships and distribution. The Island Arc 3:285-308
Mitchell, E.D. 1961. A new walrus from the imperial Pliocene of Southern California: with notes on odobenid and otariid humeri. Los Angeles County Museum Contributions in Science 44:1-28
Mitchell, E.D. 1968. The Mio-Pliocene pinniped Imagotaria. Journal of the Fisheries Research Board of Canada 25(9):1843-1900.
Mitchell, E.D. and R. H. Tedford. 1973. The Enaliarctinae: A new group of extinct aquatic Carnivora and a consideration of the origin of the otariidae. Bulletin of the American Museum of Natural History 151(3):203-284
Repenning, C.A. and R. H. Tedford. 1977. Otarioid seals of the Neogene. Geological Survey Professional Paper 992:1-93
Takeyama, K. and T. Ozawa. 1984. A new Miocene otarioid seal from Japan. Proceedings of the Japan Academy Series B Physical and Biological Sciences 40(3):36-39
1 comment:
This promises to be an interesting series of posts! Thanks for making a summary about this interesting evolutionary story that is completely ignored in popular works.
Post a Comment